
13776 работ.
A B C D E F G H I J K L M N O P Q R S T U V W X Y Z Без автора
Ученые креационисты отвечают своим критикам
Часть II
Глава 7, «Молекулярные свидетельства в пользу эволюции», написана Томасом Х.Джуксом, биохимиком, профессором биофизики в калифорнийском университете, Беркли. Автор этой главы — убежденный антикреационист. Вся она содержит в основном стандартные эволюционистские объяснения молекулярных сходств и различий растений и животных, взятые из учебников. Многие эволюционисты, отчаявшись найти свидетельства в пользу эволюции в ископаемых останках, переключились на молекулярную биологию, провозгласив, что ископаемые переходные формы нам больше не нужны, потому что молекулярная биология и так доказывает истинность теории эволюции. Обращаясь к окаменелостям, мы пытаемся понять, как возникли живые организмы в очень далеком прошлом, исследуя безмолвные свидетельства их останков. Мы рады, что наши выводы подтверждаются этими данными, но никогда не можем быть абсолютно в них уверены. Обращаясь же к молекулярной биологии, мы тоже пытаемся выяснить, что произошло в далеком прошлом, но положение наше куда сложнее, чем когда мы изучаем окаменелости. У нас нет ископаемых молекул, мы не можем вернуться в прошлое и, таким образом, анализируем гипотетические молекулы, которые тогда предположительно существовали. Единственный материал для изучения, нам доступный, — молекулы ныне живущих организмов. Мы делаем предположение и смотрим, как согласуются с ними данные молекулярной биологии, а если они не согласуются, мы изменяем свои предположения, добиваясь соответствия. Так можно оперировать данными и с креационной, и с эволюционистской точки зрения.
Давайте прежде всего рассмотрим, что общего между данными, которые ожидают увидеть креационист и эволюционист. Эволюционист ожидает, что все живые организмы будут обнаруживать некоторое сходство в биохимическом строении, — так, он считает, что жизнь возникла раз и навсегда. Первобытные протеины, ДНК, РНК и другие макромолекулы, вероятно, существовали в первоначальных формах жизни. Наряду с постепенным эволюционным преобразованием низших форм жизни в более высшие, в конечном итоге достигающим высшей точки развития в человеке, постепенно изменялись и преобразовывались и эти молекулы. Таким образом, множество или даже большинство макромолекул, обнаруженных в человеке, происходят от его далеких предков, что можно сказать и о сходных молекулах, найденных в других живых существах. В процессе эволюционного развития живых организмов эти макромолекулы претерпевают эволюционные изменения путем накопления мутаций. Различия этих молекул, протеинов, ДНК, РНК должны быть малы у близких организмов и велики — у организмов, далеких друг от друга. Так, различия аминокислотных цепочек цитохрома С человека и шимпанзе должны быть меньше, чем тех же цепочек у человека и рептилии, человека и рыбы, и т.д.
Креационист тоже будет ожидать большого биохимического сходства всех живых организмов. Мы все пьем одну и ту же воду, дышим одним и тем же воздухом и едим одинаковую пищу. Предположим, что Бог создал бы растения с одним типом аминокислот, сахара, пурина, пиримидина и т.д., потом создал бы животных с другим видом аминокислот, сахара, пурина, пиримидина и т.д., и, наконец, создал бы человека с третьим типом аминокислот, сахара и т.д. Что мы стали бы есть? Мы не могли бы есть растения, мы не могли бы есть животных; оставалось бы питаться друг другом! Очевидно, так бы дело не пошло. Все основные молекулы в организмах растений, животных и человека должны быть одинаковыми. Метаболизм растений, животных и человека, основанный на сходных биохимических принципах, должен был быть подобным, поэтому и в ключевых метаболических процессах должны были участвовать похожие макромолекулы, созданные таким образом, чтобы соответствовать специфической внутренней среде организма или клетки, в которой они должны функционировать.
Креационисты тоже полагают, что внешнее сходство существ влечет за собой и сходство молекулярного строения, и наоборот. Мы знаем, например, что человек больше похож на шимпанзе, чем на летучую мышь, но ближе к летучей мыши, чем к крокодилу или мухе. Человек, шимпанзе и летучая мышь — млекопитающие. Поэтому креационист ожидает, что его белки, ДНК и РНК — важнейшие макромолекулы, участвующие в обмене веществ — скорее подобны таким же молекулам у шимпанзе и летучей мыши, чем у крокодила или мухи. Эволюционист же считает, что эти различия возникли в результате мутаций молекул ДНК, или генов, и что они явились основной причиной различий строения растений и животных. Он считает, что все гены в живых организмах развились из ранее существовавших генов путем мутации или дупликации генов и мутаций. Креационисты полагают, что все нормальные гены, отвечающие за естественные, здоровые отличительные признаки растений и животных, были сотворены. Каждый основной тип растений или животных был сотворен с достаточным генетическим потенциалом или набором генов (генофонд, как говорят генетики), позволяющим свойствам варьироваться в границах вида, в целях приспособления к разным условиям окружающей среды.
Используя в качестве примера гемоглобин, Джукс указывает, что альфа- и бета-цепочки человеческого гемоглобина отличаются от цепочек гемоглобина шимпанзе лишь одной аминокислотой. Гемоглобин человека и крупного рогатого скота отличается 17 аминокислотами в альфа-цепочке и 24 — в бета-цепочке, и т. д. В самом деле, если накопить побольше информации, можно составить «фамильные древа» живых существ, опираясь на различия в строении гемоглобина разных организмов. Эти расхождения, пишет Джукс, совпадают со схемой «родства» живых существ, принятой согласно классической классификации животных и растений (с. 119).
Если предположить, как объясняется далее, что мутации происходили регулярно, постепенно накапливаясь, то аминокислотные различия одноименных белков, или нуклеотидные отклонения в генах, можно использовать в качестве «молекулярных часов», которые показывали бы время возникновения разнообразных животных в истории эволюции. Конечно, сразу же возникает множество побочных вопросов. Что взять за точку отсчета времени? Каковы будут «деления» часов? Определяется это с помощью предположения о том, когда определенный организм возник впервые или когда два организма разошлись в своем строении. Например, если предположить, что два организма разошлись в строении 600 млн. лет назад и что их альфа-цепочки гемоглобина различны на 60%, получается, что мутации альфа-цепочки гемоглобина идут со скоростью 1% в 10 млн. лет. Итак, если два других организма имеют различия в строении гемоглобинной цепочки 10%, получается, что они «разошлись» 100 млн. лет назад. Конечно, точность зависит в первую очеред!» от точности предположения, которое задает размер деления часов (600 млн. лет). А оно может быть верным. Но может и не быть.
Идея использования различий белков и ДНК (они кодируют строение белков — следовательно, различия в строении белков должны быть параллельны различиям в строении соответствующих генов) для построения эволюционных филогенетических схем и создания молекулярных «часов» породила очень много проблем. Было обнаружено, что различия между разными классами белков огромны. Так, если гистоны отличаются незначительно и изменения в них в 1% соответствуют 600 млн. лет развития, то для цитохрома С время, необходимое для возникновения отклонения в 1%, было оценено в 20 млн. лет, для гемоглобина — в 5,8 млн. лет, а для фибринопептидов — всего в 1,1 млн. лет — почти в 600 раз меньше, чем для гистонов. Таким образом, все часы показывают разное время.
Кроме того, филогенетическое древо, построенное на основе наблюдений за одним классом белков, скажем, цитохромом С, может значительно отличаться от другого древа, выстроенного, например, на базе исследований инсулина, миоглобина и т.д. Винсент Демулен писал:
«Кто может серьезно относиться к филогенезу позвоночных, которые, согласно сравнению состава миоглобина, произошли от одних видов, а согласно сравнению состава гемоглобина — от других? Картина, выстроенная по данным миоглобина, разительно отличается от той, которую дает гемоглобин... Основная проблема здесь — достоверность эволюционных реконструкций, основанных на данных строения цепочек... Сравнение отдельных эволюционных схем выявляет всю их слабость»[15].
Когда было проведено сравнение цитохрона и несерных пурпурных фотосинтезирующих бактерий, оказалось, что полученные данные противоречат предсказаниям эволюционистов. Т.Э.Мейер, et al, сообщает:
«Результаты наших исследований показали, что получить непосредственную филогенетическую информацию из строения цепочек протеинов родственных бактерий не так-то просто... У подобных видов различия в строении цепочки цитохрома С-551 не меньше, чем у митохондриальных млекопитающих и насекомых, хотя оба класса белков, вероятно, обладают сходными окислительно-восстановительными функциями»[16].
После исследования некоторых других цитохромов С в этих бактериях Р.П.Эмблер, Т.Е.Мэйер и М.Д-Кэмен заключили:
«...филогенетические древа, построенные в результате опытов и показанные в таблице 1, нельзя считать бесспорными и всецело соответствующими истине»[17].
Сложности с цитохромом С наблюдаются не только у бактерий. Так, когда сравнивают цитохромы С двух близких организмов, Desulfovibrio vulearis оказалось что, несмотря на сходство молекулярного веса, длины цепочки объемов отдельных частей и количества генов, они заметно отличаются по составу аминокислот[18]. Мы найдем много подобных «аномалий» и среди высших организмов. Инсулин кашалота и сейвала идентичен инсулину собаки и свиньи, но отличен от того же белка морского кита[19]. Инсулин морской свинки уникален, он значительно отличается от всех других инсулинов. Например, при сравнении аминокислотной цепочки инсулина морской свинки с инсулином человека и крысы, ее собрата-грызуна, в обоих случаях оказывается 18 различий[20]. Интересно, как бы выглядело филогенетическое древо, построенное на основании данных исследования инсулина?
Структура цитохрома С гремучей змеи имеет 22 отличия от цитохрома С черепахи, другой рептилии, но лишь 14 — от цитохрома С человека[21]. Аминокислотная цепочка белка яйца эмбденского гуся, лизоцима, не имеет ничего общего (а если имеет, то мало) с цепочкой белка куриного яйца[22].
Руководствуясь данными аминокислотных цепочек, большинство эволюционистов заявляют, что ближайший родственник человека среди человекообразных обезьян — шимпанзе, сразу за которым следует горилла. Орангутан более «удален» от человека, чем гиббон, за которым следуют прочие обезьяны. Так, по результатам экспериментов с ДНК, процентные различия в ее строении между человеком и другими приматами таковы: шимпанзе — 1,6; горилла — 2,2; орангутан — 3,7; гиббон — 5,2; бабуин — 7,4; паукообразная обезьяна — 12,6. В эволюционистских кругах эти данные широко признаны как доказательство того, что ближайший родственник человека — шимпанзе. Дж. Х.Шварц, однако, с этим не согласен. Говоря о молекулярных данных, он пишет:
«...общее сходство вовсе не обязательно и не может быть основанием для выводов о близком родстве»[23].
Креационисты согласны с ним в том, что подобие не обязательно указывает на родство как на происхождение от общего предка. На основании морфологических черт сходства и различия Шварц доказывает, что по строению человек ближе к орангутану, чем к африканским человекообразным обезьянам (шимпанзе и горилле).
Данные радиоиммунопроб и хроматографического анализа гипоталамического лютеинизйрующего гормона гормонной секреции (ЛГГС) показывают, что у млекопитающих и амфибий он неразличим, но он химически отличается от того же гормона рептилий, птиц и телеостеанских рыб. Конечно, на всех эволюционных филогенетических схемах люди и другие млекопитающие представлены как прямые потомки рептилий, вместе с птицами, а рептилии — как потомки амфибий. По этим схемам, амфибии появились раньше, чем млекопитающие, птицы и рептилии, но молекулярные данные свидетельствуют об обратном. Это побудило Дж. Э. Кинга к следующему утверждению:
«Обнаружение того, что ЛГГС амфибий идентичен пептиду млекопитающих и отличен от ЛГГС птиц, рептилий и телеостеанских рыб, которые неотличимы друг от друга при наших методах исследования, подтверждает современную филогенетическую схему, предполагающую, что млекопитающие и амфибии могут быть более родственными друг другу, чем рептилиям и птицам»[24].
В научной литературе можно найти много подобных «аномалий». Если бы действительно можно было на основе данных аминокислотных цепочек и сходствах и различиях ДНК выявлять эволюционное родство и составлять филогенетическое древо, это древо было бы единственным, и существовала бы только одна подобная схема, совпадающая со схемой, созданной по морфологическим признакам. Мы же, напротив, видим, что на основе сравнения молекулярных данных выявить одно, полностью достоверное филогенетическое древо невозможно. Более того: если показатели, полученные при рассмотрении одного протеина, использовать для построения филогенетической схемы, неизбежно не только появление аномалий, но придется «приглаживать» данные даже для доказательства очевидной родственности или подобия. Так, для того, чтобы выявить похожие части аминокислотной цепочки белка, необходимо провести ряд гипотетических вставок, делений, изменений порядка считывания гипотетических дупликаций генов и прочих генетических «манипуляций». Другими словами, даже при работе с одним белком часть данных приходится отвергать как «аномальные», а многие другие — видоизменять, чтобы подогнать к теории.
Эволюционисты Кристиан Швабе и Грегори Уорр и некреационист Майкл Дентон выступили с критикой возможности использования данных аминокислотных цепочек для построения эволюционных филогенетических схем. Основываясь на материале своих опытов, Швабе и Уорр опровергли общепринятую догму о том, что все гены, клетки и организмы развились из одной-единственной клетки-предка. Идее монофилической эволюции они предпочли идею эволюции полифилической, постулируя, что жизнь развивалась постепенно и что основные ее формы были разделены с самого начала[25]. Итак, Швабе и Уорр считают, что все основные формы жизни образовались сразу и потом претерпели изменения; эти эволюционисты, основываясь на результатах своих исследований, согласны с креационистами в том, что данные аминокислотных цепочек белков и другая молекулярная информация не могут быть использованы для построения эволюционного филогенетического древа. Фактически, они настаивают на том, что эти данные не соответствуют монофилической теории эволюции.
Швабе и Уорр указывают, что достаточно одного доказанного несоответствия или противоречия, чтобы монофилическая интерпретация потеряла свою ценность как единственно возможное объяснение эволюции, потому что если монофилическая теория молекулярных часов истинна, значит, все отдельные филогенетические схемы, построенные на основе структуры любого из ортологических белков (инсулина, цитохрома С, миоглобина и т.д.), должны накладываться одна на другую. Швабе в особенности интересовал релаксин — гормон, отвечающий за расширение лобковой кости и размягчение шейки матки и влагалища, то есть гормон живорождения. По данным Швабе и Уорра, релаксин свиньи и крысы; так же отличаются друг от друга (55%), как релаксин акулы от релаксина и свиньи, и крысы. Судя по этим данным и предположениям, которые обычно учитывают при построении филогенетических схем на основании молекулярной информации, крыса, акула и свинья должны были появиться в одно и то же время — либо в предполагаемый момент ответвления млекопитающих (70 млн. лет назад), либо в предполагаемый момент появления акулы, которая потом стала предком млекопитающих (700 млн. лет назад). Ни один из вариантов не совпадает с тем, что принято в палеонтологии и среди эволюционистов. Подобные проблемы возникают при сравнении релаксина двух акул — песчаной тигровой и колючей. Разница в строении их белков составляет 20%; это означает, что, судя по предположениям «молекулярных» эволюционистов, они «разветвились» 20 млн. лет назад, в то время как общепринятая дата — 150 млн. лет.
Швабе и Уорр пишут и о других подобных противоречиях, основанных на данных белков. Они вынуждены отмежеваться от общепринятой теории разветвленного эволюционного филогенетического древа, созданного на основе молекулярных данных. Они утверждают:
«...основной вывод, к которому мы ютим привлечь ваше внимание, состоит в следующем: наши находки позволяю! предположить, что многие гены, образовавшиеся в результате генной дупликаций, появились очень рано в процессе развития жизни, фактически, мы можем задать вопрос, все ли гены появились в результате дупликаций, или, возможяо, все они существовали с самого начала как потенциал для развития видов» (с. 476 их статьи).
Эти выводы поразительно похожи на выводы креационистов. Различие, разумеется в том, что Швабе к Уорр верят: все живые организмы, носители генов, каким-то образом появились в результате эволюционных процессов. Креационисты перефразировали бы их заявление так:
«Данные свидетельствуют, что ни один ген не возник в результате дупликаций и что все они существовали с самого вачала, поддерживая целостность свойств каждого основного типа животных я растений и предоставляя генетический материал для возможных вариаций в рамках каждого вида».
Швабе и Уорр сравнивают общепринятое толкование, основанное на разветвляющемся филогенетическом эволюционном древе, соответствующем предпосылкам молекулярной эюлюции (рис. 1), и свое собственное толкование тех же данных (рис. 2).
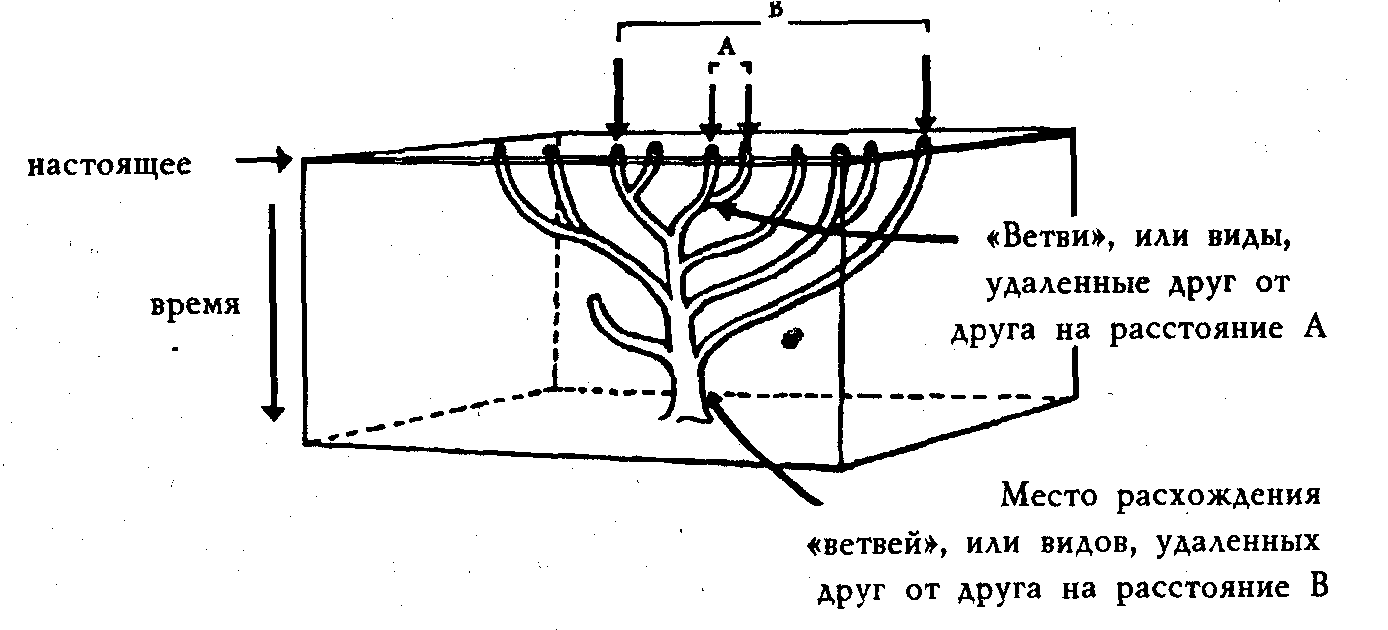
Рисунок 1. Стандартное монофилическое эволюционное древо.
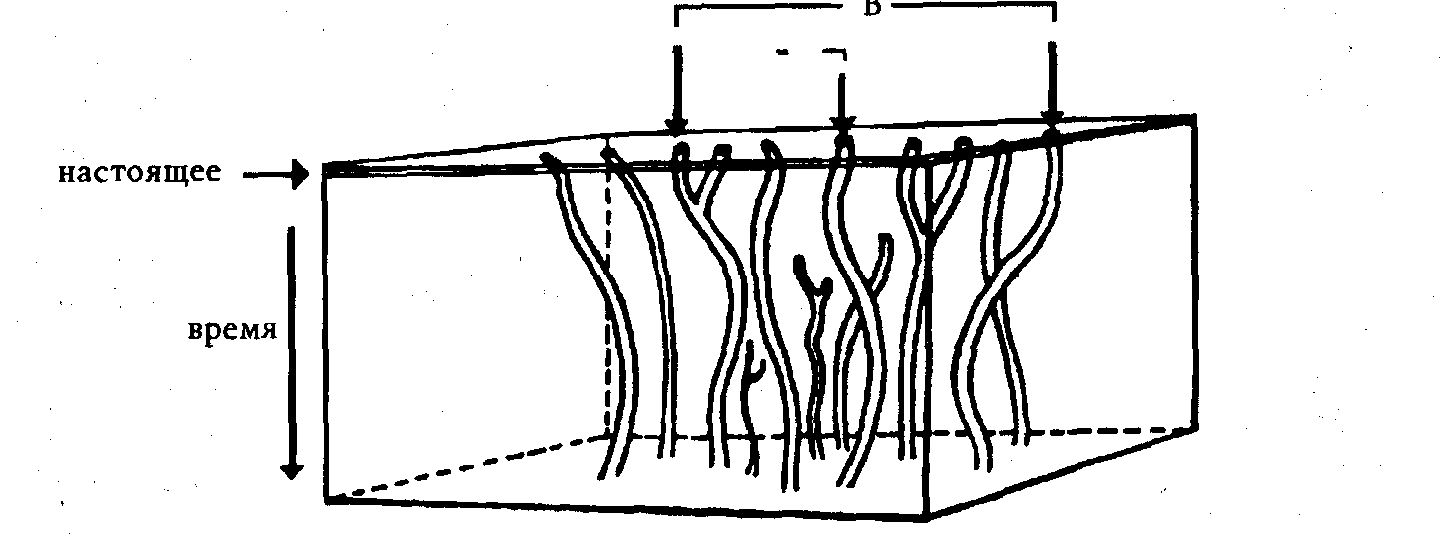
Рисунок 2. Полифилическое толкование молекулярных данных по Швабе и Уорру.
Швабе и Уорр указывают, что, если отбросить идею монофилической эволюции, любые выводы о родстве молекул или видов станут сомнительными, потому что мы не будем знать, сравниваются ли ветви одного дерева или похожие ветви разных деревьев. Если отбросить монофилическую версию, надо отказаться к от мнения о том, что сходство автоматически предполагает родство и происхождение от общего предка. Креационисты всецело согласны со Швабе и Уорром в этом вопросе.
Надо подчеркнуть, что единственные реальные данные, которые у нас есть, — это кончики веток; все остальное — спекуляции, основанные на предвзятых идеях. Исходя из молекулярных различий белков и ДНК в живущих сейчас организмах, эволюционисты строят молекулярное эволюционное филогенетическое древо с гипотетическими ветвями. Между разнообразными гипотетическими ответвлениями, которые соответствуют различиям молекул, обычно выражаемым в процентах, и современными живыми организмами, помещенными на кончиках веток, — огромная пропасть. Многие неискушенные читатели не подозревают, что цифры, помещенные между предполагаемыми предками современных белков, не были получены в результате анализа настоящих молекул, это — гипотетические цифры, предположения сторонников монофилической эволюции. Таким образом, доказательство эволюции — это всего лишь предположение эволюции.
Мне не раз приходилось принимать участие в дебатах с Винсентом Саричем, биохимиком и профессором факультета антропологии Калифорнийского университета, Беркли, одним из главных создателей гипотезы молекулярных часов. Сарич призывал своих слушателей как-нибудь иначе истолковать молекулярные данные, полученные им и другими эволюционистами. Джукс, говоря об эволюционистской интерпретации исследования молекул гемоглобина, тоже заявляет, что альтернативного объяснения нет. Вот что он пишет: «Эти данные важны, потому что они понятны только в рамках эволюционной идеи [разветвленного древа]» (с. 119 его главы). Так вот, теперь у нас наконец-то появилась альтернатива молекулярным идеям эволюционистов, и исходит она от их же коллег (причем вполне отвечающая взглядам креационистов)! Сарич сетует, что у Швабе и Уорра другие данные — что у них нет данных. У Швабе и Уорра, как вы обнаружите, читая их статью, очень много данных (и вполне доступных). Они не приводят цифр, как другие эволюционисты, потому что знают: цифры — фикция, ведь если разветвления не было, то и цифры учитывать нельзя. И хотя Швабе и Уорр не имели такого намерения, их работа и выводы очень помогают креационистам в истолковании молекулярных данных.
В своей книге «Эволюция: кризис одной теории»[26] в главе «Биохимическое эхо типологии» Майкл Дентон приводит факты, из которых становится ясно: молекулярная биология отнюдь не доказывает истинность эволюции. Дентон, специалист по молекулярной биологии, доктор наук, получивший ученые степени в британских университетах, генетик, проводивший исследования в Австралии, пишет:
«С останками ничего не вышло: морфологические свидетельства в лучшем случае спорны; может быть, новая область науки — сравнительная биохимия — наконец даст нам объективные доказательства связей, о которых так давно мечтают биологи-эволюционисты.
Однако по мере того как, начиная с 1960 г., нам становится известно все больше белковых цепочек, все яснее, что молекулы не столько свидетельствуют о родстве и преемственности, видов в природе, сколько лишь вновь подтверждают традиционное представление о том, что система природы полностью подчинена высокоорганизованной иерархической схеме, в которой нет ни одного прямого указания на эволюцию. К тому же разделение оказалось математически более совершенным, чем предполагали самые суровые типологи»[27].
Дентон указывает, что при исследовании молекулярной информации цитохрома С каждый идентифицируемый подкласс цепочек изолирован и отличен от других. Каждая цепочка может быть без колебаний отнесена к определенному подклассу. То есть существует группа аминокислотных цепочек, свойственных рептилиям, такая же группа аминокислотных цепочек, свойственных млекопитающим, телеостеанским рыбам, птицам, насекомым, покрытосеменным (цветущим) растениям, дрожжам и т.д. Ни одна цепочка или группа цепочек не может считаться промежуточной между двумя другими. Переходные или промежуточные классы отсутствуют. Все цепочки каждого подкласса в равной мере изолированы от членов другой группы.
Итак, как указывают процентные различия цепочек, цитохром С бактерии отличается от такого же белка лошади (млекопитающего) на 64%, голубя (птицы) — на 64%, шелкопряда (насекомого) — на 65%, пшеницы (растения) — на 66%, дрожжей (грибка) — на 69%. Значит, нет никакой разницы, млекопитающее ли перед нами, птица, насекомое, рыба или растение. Цитохром С шелкопряда отличается от такого же белка лошади на 27%, голубя — на 25%, черепахи — на 26%, карпа — на 25%, речной миноги — на 30% — вновь никакой существенной разницы. Цитохром С карпа (рыбы) отличается от цитохрома С лошади, кролика (млекопитающих), черепахи (рептилии) и лягушки-быка (амфибии) на 13% в каждом случае, а от цитохрома цыпленка (птицы) — на 14%. В случае с гемоглобином: гемоглобин речной миноги (циклостом или рыба без челюстей — предположительно одна из самых примитивных ныне существующих рыб) отличается от гемоглобина карпа (челюстной рыбы) на 75%, лягушки (земноводного) — на 81%, цыпленка (птицы) — на 78%, кенгуру (сумчатого) — на 76% и человека (плацентарного млекопитающего) — на 73%. Этот и другие примеры показывают, что здесь нет и намека на традиционные эволюционистские «серии», иначе, в последнем случае, мы получили бы следующее: циклостом — челюстная рыба — амфибия — рептилия — птицы и млекопитающие. Человек столь же близок к речной миноге, как и карп! Карп не занимает промежуточного места между речной миногой и земноводными. Амфибии не размещаются между рыбами и птицами, сумчатыми или людьми. Нет ни одной промежуточной группы. Одни и те же цифры получаются при сравнении гемоглобина улитки (брюхоногого моллюска) с гемоглобином речной миноги — 85%; карпа — 87%; лягушки — 87%; цыпленка — 85% и кенгуру — 85%. Эта столь почитаемая современными эволюционистами информация, основанная на данных аминокислотных цепочек, не позволяет классифицировать речную миногу как примитивный организм по сравнению с другими беспозвоночными или как промежуточное звено между улиткой (беспозвоночным) и любым позвоночным.
Дентон рассматривает дополнительные биохимические данные и утверждает, что в этих данных налицо систематические пробелы — не обнаружено плавного перехода от одной формы к другой, начиная с «примитивных» организмов и кончая самыми «продвинутыми», хотя на сегодняшний день сравнению подвергнуты сотни видов и тысячи различных цепочек белков и ДНК. Так как каждый класс на молекулярном уровне уникален, изолирован и не связан ни с какими промежуточными формами, Дентон напоминает нам, что молекулы, как и окаменелости, не дали нам доказательств того, что промежуточные звенья, предмет усердных поисков эволюционистов со времен Дарвина, когда-либо существовали.
До того как это стало известно, многие эволюционисты предсказывали, что современные организмы, которые предположительно примитивны, будут обладать белковыми цепочками, подобными цепочкам их древних предков, следовательно, окажутся посредниками между еще более древними видами и некоторыми современными, такими, как млекопитающие. Дентон цитирует Цукеркандла[28]:
«Современные организмы, очень похожие на организмы древние, вероятно, содержат полипептидные цепочки [белков], во многом схожие с цепочками этих древних организмов. Другими словами, определенные животные являются как бы "живыми окаменелостями", это таракан, подковоносы, Краб, акула, и, среди млекопитающих, лемур. Они, вероятно, вырабатывают большое количество полипептидных молекул, почти не отличающихся от тех, которые вырабатывали их предки миллионы лет назад»[29].
Упомянув, что это мнение противоречиво, Цукеркандл, тем не менее, предпочитает его. Эволюционисты часто заявляют, что любой прогресс в научном знании, особенно в биологии, полезен для теории эволюции. Далекое от истины заявление — сведения сравнительной молекулярной биологии, опровергающие предсказание Цукеркандла, лишь доказывают, что новые знания не укрепляют теорию эволюции, а добавляют трещин на ее фасаде.
Если в каждом из семейств белков — возьмем, к примеру, гемоглобин, наблюдаются существенные колебания между классами млекопитающих, птиц, рептилий, амфибий, рыб и т.д., причем ни один из этих классов, вопреки ожиданиям Цукеркандла и многих других эволюционистов, не является промежуточным, как можно называть это «аномалией»? Почему различия в строении гемоглобина миноги, карпа, лягушки, цыпленка, кенгуру и улитки одинаковы (85%)? Почему показатели гемоглобина речной миноги не являются промежуточными между показателями гемоглобина улитки и карпа, а показатели гемоглобина лягушки — карпа и цыпленка? Эволюционисты пытаются обойти эту «аномалию», доказывая, что линии, ведущие к речной миноге, карпу, лягушке и цыпленку ответвились от ствола беспозвоночных (улитка — это, конечно, беспозвоночное), скажем, около 500 млн. дет тому назад; молекулярная эволюция каждого из этих организмов насчитывает 500 млн. лет, поэтому неудивительно, что показания степени «удаления» каждой из этих групп от улитки по гемоглобину, цитохрому С и другим белкам равны, как указывают на то процентные соотношения. Это уже упоминавшаяся нами идея «молекулярных часов» — представление о том, что темп мутаций генов, кодирующих строение каждой группы белков всех животных, какими бы разными они ни были, одинаков. Так, эволюционисты считают, что во всех организмах происходило изменение состава гистонов на 1% каждые 600 млн. лет, цитохрома С — каждые 20 млн. лет, гемоглобина — каждые 5,8 млн. лет, а фибринопептидов — каждые 1,1 млн. лет. Эволюционисты верят, что эволюционные молекулярные часы разных групп белков шли с разной скоростью, то есть что у нас есть сотни часов, идущих с разной скоростью.
Эволюционисты верят, что существуют какие-то неизвестные нам факторы, заставляющие гены разных видов белков развиваться с равной скоростью.
У эволюционистов нет никаких эмпирических данных, с помощью которых они могли бы объяснить, почему гены одного вида белка развиваются быстрее, чем другого. Этого эволюционисты не предвидели и не ожидали. Более того: почему гены одного вида белка, скажем, цитохрома С, развиваются одинаковыми темпами во всех организмах, независимо от того, «примитивные» они или «продвинутые», на протяжении сотен миллионов лет? Амфибии предположительно образовались 400 — 450 млн. лет назад. С тех пор они не слишком изменились — сегодня амфибии не многим отличаются от своих представленных окаменелостями предков. Ответвившаяся от амфибий линия, наоборот, очень сильно изменилась и через рептилий и птиц привела к млекопитающим. Предполагается, что животные развивались и изменялись, потому что развивались их гены. Если гены цитохрома С, гемоглобина и других видов белков за прошедшие 500 млн. лет развивались в амфибиях так же, как и в белках других организмов, почему тогда амфибии замерли в своем развитии, а другая линия привела к появлению человека?
Насекомое совсем не похоже на млекопитающее. Если гены насекомого развивались или подвергались мутациям в том же темпе, что и гены той линии, которая привела к млекопитающим через сотни миллионов лет после того, как насекомое стало насекомым, почему насекомое до сих пор остается насекомым, а линия, ответвившаяся от него, в своем развитии дошла до человека? Как могло так случиться: в генах, задающих код белка, произошел один и тот же процент изменений, но летучие драконы, тараканы, сороконожки, пауки, клещи, кузнечики и т.д. выглядят почти так же, как, предположительно, выглядели и 350 млн. лет назад?
Кроме того, степень мутаций измеряется в количестве мутации на один ген на одно поколение, и наблюдаемый показатель примерно одинаков для многих генов. Для более высоко организованных существ степень мутации равна приблизительно 106 на ген в поколение. Другими словами, каждый ген в одной особи из миллиона в каждом поколении подвергается какой-то мутации — в лучшую сторону, в худшую или нейтральной. В таких видах, как человек и шимпанзе, длительность жизни которых не намного отличается, следовало бы ожидать похожих темпов мутации. А у мышей, напротив, за год сменяются 4 — 5 поколений. Слон достигает половой зрелости в 14 лет; его период беременности равен 18 — 21 месяцам. Получается, что темп смены поколений у мыши в 100 раз выше, чем у слона, но белок мыши так же отличается (в процентах) от белка, скажем, карпа (или любого другого «низшего» существа), как и белок слона.
Еще более резко отличаются сроки смены поколений у насекомых. Фруктовая муха производит на свет потомство за 2 недели, а цикаде для воспроизведения нового поколения требуется 17 лет. Таким образом, темп смены поколений фруктовой мухи в 1000 раз выше, чем у цикады. Если, как верят эволюционисты, современные порядки насекомых возникли около 50 млн. лет назад, за это время сменило друг друга в 50 миллиардов раз больше поколений фруктовой мухи, чем поколений цикады, но белки всех порядков насекомых, включая фруктовую муху и цикаду, одинаково отличаются от белков беспозвоночных, бактерий, растений, грибов и т.д. Если же рассмотреть такие организмы, как деревья и бактерии, разница в сроках смены поколений возрастет до 10.000 раз.
Каким чудом акула, сотни миллионов лет подвергаясь мутациям, все же осталась акулой, а какие-то другие рыбы тем временем превратились сперва в амфибий, потом в рептилий, потом в млекопитающих и, наконец, в человека? Если считается, что темп мутаций у всех животных одинаков (именно на этом основана идея о молекулярных часах), а степень мутации измеряется в количестве мутаций на ген на поколение, то почему белки организмов с огромнейшим разрывом в темпах смены поколений все-таки в равной мере отличаются от предполагаемых предков?
Как вычислили эволюционисты, скорость мутаций гистона составляет 1% в 600 млн. лет, а гемоглобина — 1% в 5,8 млн. лет — более, чем 100-кратная разница. Почему все виды гемоглобина во всех различных филогенетических линиях, будь то муха, улитка, лягушка, тунец, канарейка, корова, крокодил или человек, развиваются ровно в 100 раз быстрее, чем все виды гистона и тоже во всех филогенетических линиях? Если кто-нибудь сможет составить схему для «объяснения» этой загадки, ему придется составлять и другие схемы для объяснения систематических различий темпов мутаций цитохрома С и гемоглобинов, гемоглобинов и миоглобинов, гемоглобинов и разных протеаз, ad infinitum. Как могли естественные процессы, включающие случайные генетические ошибки, предположительно зависящие от условий окружающей среды, таких различных в разных частях света и по-разному действующих на столь многообразные организмы, привести к равномерным темпам мутаций гемоглобина во всех живых существах, и другой, но не менее равномерной скорости мутации цитохрома С во всех существах?
Завершая главу на эту тему, Дентон справедливо замечает:
«Хотя никто не может убедительно объяснить, как случайные эволюционные процессы привели к таким упорядоченным различиям, идея всеобщности сроков эволюции представлена в литературе как доказанная опытным путем. Догма эволюции столь влиятельна, что идея, больше похожая на принцип средневековой астрологии, чем на серьезную научную теорию XX века, для биологов-эволюционистов стала реальностью»[30].
Теперь, когда мы подвергли критике идеи молекулярной теории эволюции и привели характеристику Дентона, считающего эту идею сходной с принципом средневековой астрологии, о главе Джукса «Молекулярные доказательства теории эволюции» осталось добавить совсем немного. Может оказаться полезным последний комментарий. По уверению Джукса (с. 125), доказано, что прокариоты развились в эукариотов, когда некоторые из прокариотов были атакованы бактериями, которые изменили свою структуру и функции, став митохондриями, а некоторые были атакованы цианобактериями, которые в ходе подобного процесса стали хлоропластами. Этому заявлению противоречат слова коллеги Джукса, биохимика Винсента Демулена, который заявил: «Что касается происхождения эукариотических клеток, мнение о том, что есть данные, подтверждающие их происхождение в результате серии симбиозов необоснованно»[31]. Идея о том, что одни бактерии атаковали другие бактерии, или прокариоты, чтобы затем, вступив с ними в отношения симбиоза, превратиться в митохондрии или хлоропласты, обычно носит название теории о симбиотическом происхождении эукариотов.
Связь между прокариотами и эукариотами отрицает и Джеймс Дарнелл. Основываясь на опыте своих исследований, Дарнелл провозглашает:
«Биохимические различия информационных РНК, формирующихся в эукариотах и прокариотах, столь глубоки, что эволюция от прокариотической клетки к эукариотической представляется невероятной»[32].
Вот чего стоит «доказано» Джукса!
Глава 8, «Недавние похороны Дарвина — в который раз!» (с. 139-146), написана Стивеном Джеем Гоулдом и представляет собой переиздание очерка Гоулда в «Нэйчурал хистори» 1976 г[33]. Этот очерк был посвящен критике статьи Тома Бетелла в «Харперз Мэгэзин»[34], в которой Бетелл выдвигает тезис о том, что дарвинистская теория эволюции, основанная на естественном отборе, рушится, потому что стало ясно: естественный отбор — не более, чем тавтология. Так как критику Бетелла и контраргументы Гоулда мы уже рассмотрели в главе 3 этой книги, нет необходимости еще раз обсуждать главу Гоулда. Напомним лишь, что, судя по словам философа Рональда Брэди, попытка Гоулда спасти статус естественного отбора как научной теории с треском провалилась[35].
Глава 9, «Теологические и палеонтологические аргументы, креационизма» (с. 147-162), написана Дейвидом М.Раупом. Бывший декан по научной работе Полевого музея естественной истории, Рауп теперь занимает должность председателя отделения геофизических наук Университета Чикаго. Он широко известен, благодаря своим исследованиям в палеонтологии беспозвоночных и геологии, является членом Академии наук. Вместе со Стивеном М.Стэнли он написал учебник «Введение в палеонтологию»[36]. Читать главу Раупа — великое облегчение, ибо, в отличие от многих других глав сборника, здесь нет невыдержанной полемики с креационистами и креационизмом. Не оставляет сомнений то, что Рауп верит в эволюцию, но к противоречию эволюционизм/креационизм он подходит спокойно и объективно. Высказывания Раупа резко контрастируют со словами Гоулда, неучтиво называющего креационистов «йеху», и заявлениями других коллег Раупа — эволюционистов, утверждающих, что креационизм не похож на науку и есть не что иное, как замаскированная религия: Рауп считает, что методы креационистов вполне научны (хотя иногда, по его мнению, не в той мере, в какой хотелось бы) и что статус и ценность креационизма не зависят от идеологии или религиозных верований ученых-креационистов. Итак, Рауп утверждает:
«Часто говорят, что креационисты интересуются только идеологией (библейской) и недостаточно свободны интеллектуально, чтобы рассматривать проблемы происхождения жизни в приемлемой научной манере. Не секрет, что между поддержкой креационных идей и исповеданием религии существует связь. Но все чаще люди, не имеющие твердых религиозных убеждений, вслушиваются в доводы ученых-креационистов и принимают их. Таким образом, идеология в какой-то степени играет роль, но важна не только она. Кроме того, я считаю, что вопрос о том, научны аргументы или нет, следует решать независимо от идеологии тех, кто их выдвигает».
Ученые-креационисты, я думаю, согласятся с таким заявлением. Каковы бы ни были идеология и религиозные верования эволюционистов и креационистов, их научные теории по поводу происхождения жизни следует оценивать независимо от этих верований, основываясь только на научных доказательствах, подтверждающих их взгляды. Но, конечно, лишь очень немногие люди непредвзято относятся к вопросу о происхождении жизни.
Рауп подтверждает, что креационисты прибегают к научным методам и подвергают проверке идею сотворения. Он пишет:
«Было опубликовано несколько креационных схем-гипотез (Гиш, 1978, с. 50-51, например). Проверка этих гипотез часто включает в себя очень тщательно разработанные, а иногда — удивительно результативные эксперименты. Недавний пример тому — переоценка широко известных окаменелых отложений силурийской системы из Торнтон Квэрри, Иллинойс (Д'Армонд, 1980). Эти останки считались остатками погребенных рифов, и многие годы проводились работы по их изучению. Д'Армонд предположил, что эти осадочные породы — результат грандиозного потопа, и, хоть я и не согласен с его анализом и выводами, это исследование — очевидная попытка использовать геологические данные для подтверждения креационной модели.
Итак, хотя ученые-креационисты твердо верят в авторитет Библии, они не пытаются вести исследование, полагаясь только на ее авторитет. Напротив, во всевозможных источниках они ищут подтверждающие это исследование данные. Теоретически, креационист вроде Д'Армонда мог бы отвергнуть креационную модель, не обнаружив подтверждающих ее геологических данных. Это совсем не похоже на ревностного креациониста, но научная креационная литература дает заинтересованному непрофессионалу основание заключить, что библейское повествование подгоняется к научным данным. На самом деле, креационисты подвергают библейское повествование о сотворении научной проверке».
Хотя Рауп и считает, что в труды креационистов просочились фактические ошибки и недопонимание (такое можно сказать, конечно, и о работе ученых в целом), он пишет:
«По моему мнению, немногие из аргументов, используемых креационистами, "научны" в том плане, что они основываются на базовых методах проверки гипотез, обычно считающихся научными (с. 159)... некоторые из ученых-креационистов потрудились на славу, проникая в сложнейшую область науки...» (с. 160).
Основная задача Раупа как одного из авторов указанного сборника, по его собственным словам, — это критика геологических и палеонтологических доказательств креационистов, и он очень добросовестно подходит к решению этой задачи. Так, он говорит:
«Гиш (1978) популяризовал идею того, что скалы и ископаемые останки говорят НЕТ эволюции. Я же покажу здесь, как скалы и останки говорят ей ДА!» (с. 147).
Креационисты расходятся с Раупом во мнении о том, кто же победил в споре, а любой читатель главы, написанной Раупом, увидит, что его аргументы по поводу окаменелостей весьма и весьма слабы.
Рауп начинает спор с искреннего признания: с эволюционистской точки зрения, ископаемых останков у нас очень мало. Он пишет (с. 156):
«Дарвин предсказал, что ископаемые останки должны являть гладкую преемственность, плавные переходы от предков к потомкам, и содержать удовлетворительное число промежуточных звеньев между основными группами. Дарвин даже заявил, что, если результаты раскопок окажутся иными, его теории эволюции будет угрожать серьезная опасность. Во времена Дарвина таких плавных переходов обнаружено не было; он объяснял это недостаточным количеством палеонтологических находок и недостаточно глубоким их изучением. С тех пор прошло сто лет, и ситуация не слишком изменилась. Со времен Дарвина знания палеонтологов неизмеримо выросли, теперь мы знаем об окаменелостях значительно больше, но по сути ситуация осталась прежней. Сейчас у нас еще меньше примеров плавных переходов, чем было во времена Дарвина, потому что некоторые старые примеры при более внимательном рассмотрении оказались несостоятельными. Да, были обнаружены новые промежуточные формы, в основном среди сухопутных беспозвоночных. Но если бы Дарвин писал свою книгу сегодня, он бы все еще ссылался на смущающее отсутствие переходных форм или недостающих звеньев между крупнейшими группами организмов».
Дарвин признавал, что окаменелости могут стать сильнейшим доказательством его теории. Он надеялся и ждал, что будущие исследования откроют нам переходные формы, необходимые для подтверждения теории эволюции, и утверждал, как напоминает нам Рауп, что, если эти переходные формы не будут обнаружены, его теории эволюции (теории пути от амебы до человека) будет угрожать серьезная опасность. Со дня публикации книги Дарвина прошло 133 года, а ситуация еще хуже, чем тогда. Не только не удалось найти переходные формы, которые должны существовать, если теория эволюции — правда, но и многие из окаменелостей, которые современники Дарвина считали останками переходных форм, оказались вовсе не ими. Рауп, как многие эволюционисты, заявляет, что кое-какие переходные формы все-таки существуют, и приводит в пример археоптерикса. Пример очень спорный и, кроме того, если бы эволюция имела место, у нас было бы много тысяч бесспорных переходных форм.
Рауп обещает доказать, что окаменелости говорят эволюции ДА. Как же он это делает? Во-первых, как описано выше, он честно признает сложность проблемы окаменелостей для теории эволюции. Затем он описывает, почему переходные формы не были найдены, или, выражаясь его словами, отвечает на вопрос, «как эволюционисты объясняют недостаток промежуточных форм?» (с. 156). Для начала Рауп утверждает, что отчасти это зависит от искусственности нашей системы таксономической классификации. Например, Рауп пишет, что археоптерикс — переходная форма от рептилии к птице, таксономисты же вынуждены причислить его к тому или другому классу — поэтому животное было произвольно определено как птица. Ученые-креационисты указывают, что последние несколько лет велись анатомические исследования черт археоптерикса и было установлено, что они подобны скорее чертам птицы, чем рептилии, и что по прошествии времени его статус переходной формы становится все более сомнительным. Кроме того, если животное или растение занимает промежуточное место между двумя крупными категориями, его «промежуточность» должна бросаться в глаза, независимо от того, какое место ему отведут таксономисты. Если бы археоптерикс был покрыт наполовину чешуей, а наполовину — перьями, и его перьевой покров не был бы идентичен покрову современных птиц, если бы его передние конечности являлись чем-то средним между лапами и крыльями, а не напоминали бы птичьи крылья, было бы сразу видно, что это — промежуточная форма между рептилиями и птицами. Приведем другой пример: бы если какие-нибудь беспозвоночные, подобные трилобитам, превратились в рыб, переходные формы легко можно было бы распознать, независимо от того, определили бы их таксономисты в класс Trilobita или в класс Pisces.
Второй аргумент Раупа несколько странно слышать от такого знающего геолога, — он жалуется, на бедность окаменелостей. На это же ссылался Дарвин, но сегодня многие геологи отказались от этого довода как от не соответствующего действительности. В настоящее время мы обладаем богатейшими находками ископаемых останков — более 250.000 различных видов ископаемых классифицировано и хранится в музейных коллекциях. На с. 158 Рауп, кажется, забыл о том, что писал на с. 156, где утверждал:
«Со времен Дарвина знания палеонтологов неизмеримо расширились, теперь мы знаем об окаменелостях значительно больше...»
Так оно и есть! Палеонтологи усердно искали «недостающие звенья» по всему миру. Тщательно исследовались породы всех так называемых геологических периодов. Было сделано много новых открытий, но каждая новая находка легко находила свое место в системе порядков, классов и типов. Никаких связей между основными категориями найдено не было. Если бы теория эволюции была правдой, из 250.000 обнаруженных останков десятки тысяч, бесспорно, были бы переходными. Ссылка Раупа на бедность находок никого не обманет и не убедит.
Третье и последнее объяснение отсутствия промежуточных форм, приводимое Раупом, таково:
«Третье объяснение отсутствия промежуточных форм состоит в том, что они существовали в течение очень коротких промежутков геологического времени, если, как полагают многие теоретики, смена одного крупного типа другим происходила быстро (модель прерывистого равновесия Элдреджа и Гоулда [1972]. Это просто снижает вероятность того, что промежуточные формы будут найдены)» (с. 158).
Как указывали ученые-креационисты [37], сценарий «прерывистого равновесия» был изобретен для объяснения отсутствия переходных форм между видами, а не отсутствия переходных форм между основными типами растений и животных, такими, как семейства, порядки, классы и типы. Понятие «прерывистого равновесия» (означающее, что в результате стихийных процессов новые виды могут развиваться очень быстро без участия естественного отбора) не только не разрешает проблему пробелов между крупными категориями, но и не имеет к ней никакого отношения.
Рауп, конечно, не упоминает о непреодолимости барьеров, которые эти огромные пробелы между одноклеточными организмами и сложными беспозвоночными, а также между беспозвоночными и рыбами, ставят перед теорией эволюции. Это — самая серьезная из проблем, которые создают окаменелости для теории эволюции. Почему же Рауп ничего не говорит о ней? А просто потому, что у него, как и у других эволюционистов, нет решения. Взрывное появление полностью сформированных и очень разнообразных сложных беспозвоночных, резкое возникновение разных видов рыб — тоже полностью сформированных, без каких-либо следов предков, являются самыми точными доказательствами сотворения мира, каких только можно ожидать.
Обсуждение темы ископаемых останков Рауп завершает словами:
«Имея в виду эта соображения, мы можем утверждать, что данные раскопок совпадают с предсказаниями эволюционистов».
В статье, опубликованной Раупом два года назад, говорится:
«Дарвин, объясняя несоответствие свидетельств окаменелостей своей теории, говорил о скудости находок палеонтологов — будто бы в них полно пробелов и нам еще многое предстоит узнать. Теперь со времен Дарвина прошло 120 лет, и наши знания об окаменелостях значительно расширились. Теперь у нас есть ископаемые останки четверти миллиона видов, но ситуация не слишком изменилась»[38].
Если во времена Дарвина результаты раскопок были несовместимы с теорией Дарвина и ситуация не слишком изменилась с тех пор, то как же теперь они оказались совместимыми с теорией эволюции? Как было упомянуто выше, единственным настоящим прогрессом со времен Дарвина явилось изобретение понятия прерывистого равновесия, а это понятие, даже если оно истинно, не решило бы проблемы пробелов между высшими категориями живых организмов. Далее, если бы теория эволюции была истинной, зачем надо было бы так долго спорить, соответствуют ли ей результаты раскопок? Зачем было бы объяснять отсутствие промежуточных форм? Если бы теория эволюции была истинной, окаменелости послужили бы бесспорным ее доказательством. В начале главы Рауп заявил, что покажет, как породы и окаменелости говорят ДА эволюции. Но даже если принять все его объяснения, окаменелости все равно говорят эволюции НЕТ.
Критикуя доводы и заключения геологов-креационистов, Рауп подробно излагает взгляды этих геологов. Он объясняет, какие из их выводов считает ошибочными или устаревшими. Часть вины, признает Рауп, лежит на геологах-эволюционистах: они выдвигали ошибочные или устаревшие идеи, а креационисты зависели от их информации. Например, Рауп указывает, что креационисты слишком полагаются на факт геологической катастрофы, акцентируя возможность того, что многие большие отложения осадочных пород и обширные «кладбища» окаменелостей — последствия грандиозного всемирного потопа. В то же время, считает Рауп, креационисты приписывают геологам-эволюционистам доктрину униформизма — представление о том, что процессы настоящего, проходившие в нынешних темпах в течение длительных промежутков времени, могли привести к возникновению большей части геологических черт без вмешательства катастроф. Рауп справедливо указывает, что концепция униформизма теряет вес в глазах современных геологов. Он пишет (с. 152):
«В XIX веке сочетание геологии Лайеля и биологии Дарвина породило мудрую науку о том, что Земля и жизнь на ней развиваются постепенно, в ходе процессов, идущих одинаковыми темпами. Многочисленные примеры катастрофизма, упоминаемые теперь учеными-креационистами, были хорошо известны, но геология и палеонтология XIX века либо игнорировали их, либо придавали им второстепенное значение. С тех пор, однако, многое переменилось, и большинство современных геологов и палеонтологов принимают идею катастрофы как реальную, хоть и избегают слова "катастрофа", фактически, теперь многие геологи видят, что редкие и короткие события вносят основной вклад в формирование геологических слоев. Во многих случаях исследование "памяти" пород выявляет следы чрезвычайных событий (штормов, ураганов, землетрясений, оползней, извержений вулканов), оставивших после себя огромное количество осадочной породы, представляющей лишь краткий промежуток времени, который сбивает хронологию слоев. Периоды сравнительного затишья дают мало осадочных пород. Прошли те времена, когда геологи измеряли толщину породы и определяли возраст скалы, а потом путем деления одной цифры на другую вычисляли скорость отложения породы в сантиметрах на 1000 лет».
Рауп признает также, что отчасти в непонимании ситуации были виновны геологи-эволюционисты. Он утверждает:
«Отчасти в непонимании есть вина самих геологов: идеи униформизма и постепенности до сих пор бытуют в популярном подходе к геологии, в экспозициях некоторых музеев и текстах учебников невысокого уровня. Этому все еще учат в средней школе, так что вряд ли можно винить креационистов в том, что они говорят: традиционно принятый геологией взгляд не допускает идеи катастрофы».
Фактически, как и пишет Рауп, геологи-эволюционисты избегают слова «катастрофа», потому что всеобщее признание катастрофы событием, приведшим к зарождению жизни, подтвердило бы концепцию, которой давно придерживались креационисты, но которую консервативно отвергали эволюционистские круги. Кроме того, то, что читал Рауп из работ креационистов, уже немного устарело. Все креационисты не только знают об изменении мировосприятия геологов-эволюционистов, но рады видеть, что этот сдвиг сближает наши позиции. Геологи-эволюционисты, начиная с Джеймса Хаттона и Чарлза Лайеля в начале XIX века, исключали возможность всемирной катастрофы и ее роль в формировании облика Земли. Сейчас, однако, все большее число геологов готово уступить и признать вероятность всемирной катастрофы. Например, все еще отказываясь от принятия версии всемирного потопа (в конце концов, он ведь описан в Библии!), многие геологи склоняются к мысли, что с Землей столкнулся астероид, поднявший в воздух столько пыли, что Земля на несколько лет оказалась покрыта ею, погибли многие растения и вымерли все динозавры, исчезли некоторые другие формы жизни. Так что вот вам и всемирная катастрофа с жертвами!
Рауп выступает против заявлений некоторых креационистов, утверждающих, что построение геологических схем и датировка окаменелостей производятся на основании биологических теорий эволюционистов. Он указывает, что геологическая схема, как мы знаем, была завершена в 1815 г., почти за 50 лет до опубликования Дарвином его книги, и в значительной мере разрабатывалась креационистами. Рауп отводит существенный раздел своей главы опровержению правильности датировки пород по стадии эволюции находящихся в них окаменелостей. Рауп утверждает, что, хотя самые древние из известных нам окаменелостей — это простые прокариотические организмы, а более молодые породы содержат более сложные формы жизни, вряд ли возрастания сложности достаточно для датировки пород. Рауп считает, что процесс эволюции не прямолинеен. Однако он признает, что очень многие геологи-эволюционисты, специализирующиеся не на палеонтологии, разделяют заблуждение, на которое указывают креационисты и согласно которому геологические породы разного возраста содержат останки организмов все возрастающей сложности. Креационисты не только цитировали многие подобные высказывания из эволюционистской геологической литературы, но и на дебатах возражали тем, кто говорил, что сложность строения останков постепенно возрастает. Таким образом, Рауп вынужден был признать:
«Креационисты... ведут себя честно по отношению к этому заблуждению, хотя бы отчасти. Некоторые учителя и авторы учебников, особенно в конце XIX в. — начале XX в., были так очарованы красотой модели Дарвина, что приписывали ей могущество, которым она никогда Не обладала».
Далее Рауп говорит, что, по иронии судьбы, креационисты принимают как факт ошибочное представление о том, что геологические пласты являют нам последовательность организмов, развивающихся от простого к сложному, и стараются подогнать эти данные под модель потопа. Сейчас, когда он пояснил креационистам, что они заблуждались, он хочет искоренить - ошибку из эволюционистских учебников геологии и литературы.
Может быть, я не вполне понял, что хотел сказать Рауп, но, по-моему, он сам себе противоречит. Он пишет (с. 154): «Геохронология безоговорочно опирается на существование четкой последовательности различных предметов, находимых в породах; эта последовательность представлена, в числе прочего, окаменелостями». Но на с. 160 Рауп пишет: «Нередко, однако, сравнительно молодые породы обнаруживаются под более старыми». Конечно же, решение о том, какие породы моложе, а какие — старше, зависит от содержащихся в них окаменелостей, и если нередко более молодые породы оказываются под более старыми, то как можно утверждать, что геохронология безоговорочно основывается на «четкой последовательности»? Очень часто приходится прибегать ко вспомогательным гипотезам, чтобы объяснить, почему породы, предположительно содержащие более древние останки, находятся над породами, содержащими более молодые останки.
Заключительный раздел главы Раупа озаглавлен «Могут ли эволюционисты ошибаться?» Здесь Рауп утверждает, что эволюционисты не могут претендовать на абсолютно верное понимание истории жизни и ее процессов, и напоминает читателю о многих великих научных версиях, которые на поверку оказывались ошибочными. Он уверен, что эволюционисты правы, исчисляя возраст Земли миллиардами лет, но согласен с тем, что механизм эволюции, возможно, еще не понят. Он совершенно не понимает предложения креационистов: изучать две модели происхождения жизни. Он утверждает, что существует несколько биологических схем эволюции, то есть несколько альтернативных моделей, и креационисты неправы, говоря, что их только две: эволюционистская и креационная. Рауп не понимает того, что в рамках каждой модели, возможно, и существуют подсхемы, но основных моделей только две: натуралистическая и механистическая атеистическая теория эволюции и теистическая, сверхъестественная модель сотворения. Ламаркизм, неодарвинизм, прерывистое равновесие, подающий надежды монстр и теистическая эволюция — все это подсхемы внутри эволюционистской модели или парадигмы, в то время как прогрессивное сотворение, теория разрыва, недавнее сотворение, и т.д. — подсхемы внутри креационной модели. Эволюционист не может заявлять, что учит двум моделям о происхождении мира, если предлагает студентам альтернативные схемы эволюции.
[ Оглавление ] [ Продолжение... ]© Гребневский храм Одинцовского благочиния Московской епархии Русской Православной Церкви. Копирование материалов сайта возможно только с нашего разрешения.